Proceedings
of The World Avocado Congress III, 1995 46-51
QUANTITATIVE
APPROACH TO AVOCADO POLLINATION
Gad
Ish-Am Dan Eisikowitch
Tel
Aviv University, Department of Botany, Tel Aviv 69978, Israel
Keywords:
honey bee, Apis mellifera, Persea americana, fertilization, fruit set,
Abstract
New evidence
from Israel indicates that yield in a healthy, efficiently managed avocado
orchard, subject to no post-bloom weather-related catastrophes, is mainly a
function of honey bee activity. The early-blooming avocado cultivars were found
to be exposed to very low honey bee activity throughout most of their blooming
period, and fruit set occurred only when bee activity increased to a high
enough level. The trees along the plot's perimeter attracted higher bee activity,
resulting in a higher pollination rate, and they bore more fruit than trees
inside the plot. This effect was more pronounced when bee activity was lower.
Pollination rates of five avocado cultivars were found to be positively
correlated with honey bee activity. They averaged 60-80% pollinated stigmas
with 6-10 pollen grains per stigma on days with 20 bees per tree or more, and
only 2-10% pollination with 1-2 grains per stigma on days with less than 5 bees
per tree. Cross-pollination rates were 50-75% of the above figures on trees
adjacent to the pollen-donor cultivar, and much lower when the pollen donor was
more than two rows away.
1. Introduction
Evidence
accumulated over the last decade shows that in countries with a Mediterranean
climate honey bees are the main avocado pollinator, and that pollination may be
a limiting factor in avocado productivity (Bekey, 1989; Ish-Am &
Eisikowitch, 1991, 1993; Robinson, 1989; Vithanage, 1986). However, no
quantitative measurements have been performed of avocado fruit-set and yield
dependency on bee density and pollination rate. This work was aimed at
achieving quantitative correlations, under field conditions, between avocado
flower and honey bee densities, pollination rate, fruitlet set and yield. These
correlations could produce a tool for determining how many bees are needed to
realize the orchard's yield potential, assuming no weather-related
catastrophes.
2. Materials and methods
Observations were
carried out during the blooming seasons of 1989-1992, in avocado orchards in
the Western Galilee of Israel, both in the Kuren Valley (1-4 km from the
Mediterranean coast, 30 m above sea level) and in the hill region (10 km from
the coast, 300 m above sea level). Avocado flower and fruitlet densities were
recorded in five medium-size inflorescences per tree, over five trees per
studied cultivar. Bee density per tree (BPT) was measured on five trees per
cultivar, by counting honey bees on the whole tree for I min, while walking
around it. Pollination rate was measured using samples of 50 stigmas, which
were collected every hour from five trees per studied cultivar. The harvested
styles were placed on microscope slides coated with CMC gel containing methylene
blue and were kept at 5ºC (Melamud H., personal communication). Pollen grains
on the stigmas were later counted using a light microscope. Yield was measured
by counting fruits on 20 trees per cultivar in the autumn.
Observations
were designed to track the seasonal course of flower, bee and fruitlet
densities (three observations a week throughout the blooming season) and to
discover gradients of bee density, pollination rate and yield vs. distance from
pollen-donor cultivar and/or plot edge. Statistical calculations were performed
according to Sokal and Rohlf (1981).
3. Results
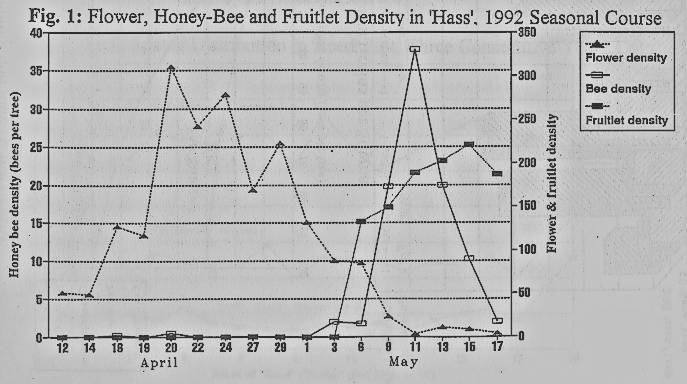
Monitoring
throughout the season of flower, honey bee and fruitlet densities of the
early-blooming avocado cultivars 'Ettinger' and 'Hass' revealed a clear
discrepancy between the seasonal courses of flowering and honey-bee activity
(Fig. 1). These cultivars received almost no bee activity throughout most of
their blooming period, including their flowering pick. Fruitlets began to
appear on the marked inflorescences only 3 days after honey bee density on the
female-stage flowers reached at least three bees per tree. This only happened
during the last 1-2 blooming weeks of these cultivars: a few fruitlets appeared
on 'Ettinger' during the first blooming week, but were too few to be counted on
the marked inflorescences.
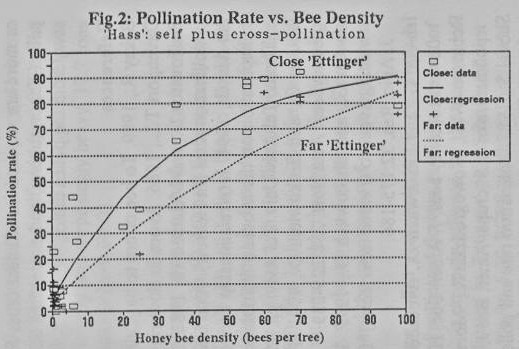
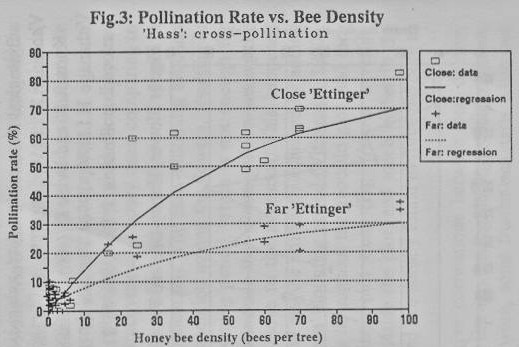
All studied
cultivars exhibited a significant positive correlation between pollination
percentage (PP) and bee density (BPT), which best fit the exponentially
ascending curve: BPT = k1*(1-Exp(-k2*BPT))+k3, where B is the function value
for BPT=0, and k1+k3 is the function's maximum asymptote (Figs. 2,3). For each
cultivar, the pollination curves yielded when a pollen-donor cultivar was
near-by were higher than those with a more distant pollen donor, though the
differences were smaller for curves of self- plus cross-pollination (at the end
of a day's bloom) and bigger for curves representing cross- pollination only.
The value of k3 was close to 0% for the cross-pollination curves and 3- 12% for
the curves of self- plus cross-pollination.
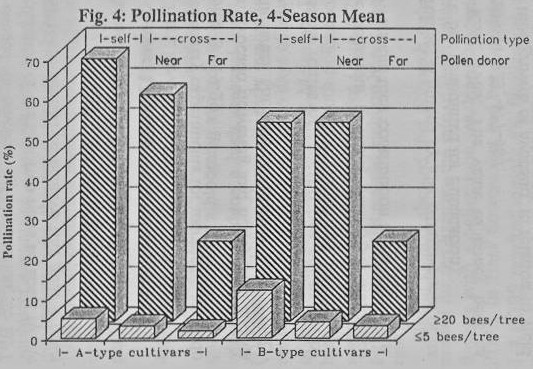
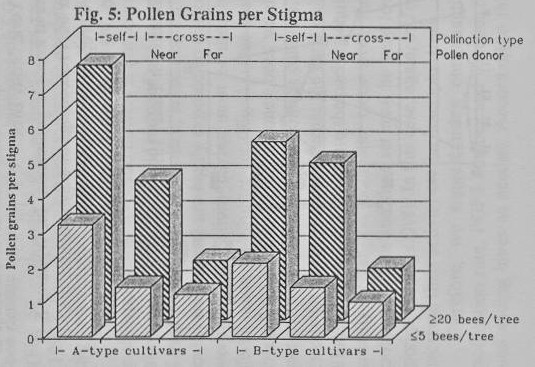
On average, when bee density
was low (5 BPT or less) the avocados achieved only 3- 10% self-pollinated
flowers with 2-3 pollen grains per stigma (PGPS), and less than 4%
cross-pollination with 1 PGPS (Figs. 4,5). However, when bee density exceeded
20 BPT, and a pollen-donor cultivar was close-by, they reached 50-70%
self-pollination with up to 7 PGPS and about 50% cross-pollination with 4 PGPS,
though only 20% cross-pollination with 1-2 PGPS when the pollen donor was more
than two rows away.
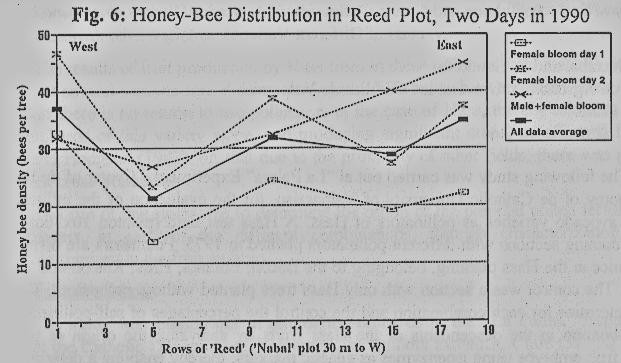
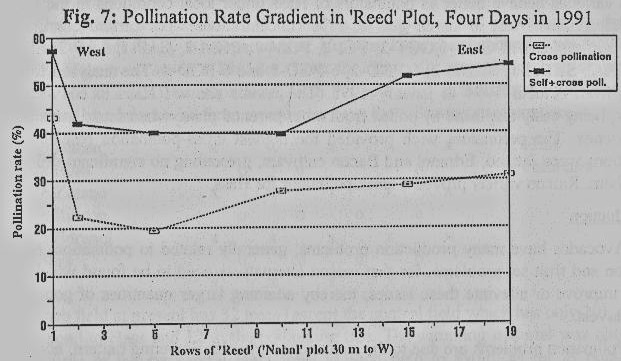
Pollination
gradient, related to distance from the plot edge, was studied in a homogeneous
'Reed' plot (19 trees by 29 rows), bordered by fallow fields on both its
northern and southern sides, a plot of early-blooming avocado cultivars to the
east (which had already completed their bloom), and a Nabal 'plot 30 m to the
west. Honey bee distribution in the plot (Fig. 6) showed a significant
preference for the perimeter trees over the trees inside the plot, with a
minimum value in the fifth tree (p=0.013, Anova on data normalized by the
maximum). Pollination rates revealed the same perimeter advantage (Fig. 7),
with minima in the fifth to tenth trees (p=0.002, Anova on the percentage
transformation).
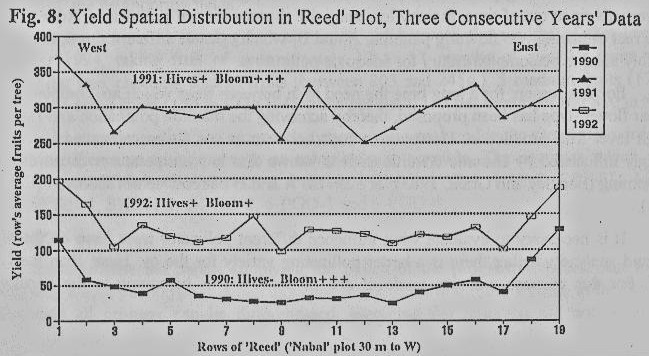
Yields
in this Reed' plot revealed a similar perimeter advantage (Fig. 8). The 1990
crop (7.4 tons per hectare) was carried almost exclusively by the perimeter
trees, which held about 18 tones per hectare, whereas the inner-plot trees were
almost empty (p=0.0001, two-way Anova). This crop resulted from a heavy bloom
the previous year, coupled with very low honey bee activity (bee hives had not
been stationed in the orchard), and the bees that did visit the heavy bloom
were seen almost exclusively on the plot's perimeter trees. In 1990 the bloom
was again very heavy, but this year bee hives were stationed in the orchard, resulting in a
high bee density and a decrease in the perimeter advantage. The following year
(1991) production was very heavy, 41.4 tons per hectare, with the
perimeter-advantage effect being less significant (p=0.002). The 1991 bloom,
occurring simultaneously with such a heavy crop, was very sparse, but was
intensively visited by the bees. The next year, the crop was surprisingly
heavy, 17.7 tons per hectare, with a clear effect of perimeter advantage
(p=0.0002).
4. Discussion
The role of
pollination as a limiting factor in avocado productivity has been questioned,
since a blooming avocado tree carries some 50 000 new flowers every day, which
could produce a high seasonal yield if only 1% of I day's bloom would set
(Davenport, 1986). However, early qualitative observations in Israel found that
fruitlet set in the field does not occur on days of low bee activity, with a
pollination percentage of less than 20% (Eisikowitch & Melamud, 1982). The
high positive correlations found here between bee density and both pollination
rate and yield, throughout the blooming season (Fig. 1) and across the plot
(Figs. 6,7,8), emphasize the dependency of avocado pollination, fruitlet set
and yield on honey bees, and the importance of a high pollination rate for
fruit set.
Shoval (1987)
showed that although only one pollen grain penetrating the ovary is needed for
fertilization, the rate of fertilization of pollinated flowers (actually, the
rate of flowers with a pollen tube reaching the ovary) is strongly affected by
the number of PGPS. In 'Hass' flowers that had been hand-pollinated by
'Ettinger' pollen, he found that 1-4 PGPS resulted in only 4% fertilization,
and even 5-19 PGPS produced only 11% fertilization. Only at 20-30 PGPS was a
substantial fertilization rate ensured. We found a significant positive
correlation between pollination percentage and PGPS, in which high bee density
corresponded to averages of 5-7 self-grains, and 4 cross-grains per stigma
(Fig. 5), with only 24% of the stigmas carrying 20 or more pollen grains (Ish-Am,
1994). Therefore, we assume that the need for both high bee density
and pollination rate for fertilization is a substantial feature of the
avocado's reproductive system.
These conclusions
may provide insight into situations of low yield caused by low pollination
rates. Avocado flowers are of low attractiveness to honey bees, comparing to
flowers of Citrus spp., wild mustard and some other cultivated, as well
as wild species in Israel (Ish-Am & Eisikowitch, submitted for
publication). The early-blooming avocado cultivars in Israel are highly exposed
to this competition among the flowers, resulting in very low bee activity
during most of their blooming period, which is not enough for fertilization and
fruitlet set (Fig. 1). One may conclude that if competition among the bees
would increase more honey bees would move to the avocados. This happened
naturally with Israel's late-blooming cultivars, and may be artificially
achieved by adding bee hives to the orchards during the early cultivars'
blooming period (Vithanage, 1990). Honey bees do not distribute themselves
equally throughout the avocado plot, and they usually prefer the perimeter
trees over the inner-plot trees (Fig. 6). As a result, pollination rate and
fruit set may be very low inside the plot, and much higher in the perimeter
(Figs. 7,8). In this case, again, increased bee density found to force the bees
to increase their activity on the inner-plot trees.
Therefore, we suggest using
our quantitative correlations between bee density, pollination rate, PGPS
number and fruit set for bee-monitoring during avocado bloom, and adding hives
as needed. According to this work, some fruitlet set starts when bee density reaches 3 BPT, but
10-20 BPT are needed for efficient self-fertilization, 25 BPT or more are
required for efficient cross-fertilization of the first two rows adjacent to
the pollen-donor cultivar, and more than 55 BPT are needed to achieve this goal
4 rows away from the pollen donor.
References
Bekey
R., 1989. To bee or not to be. Calif. Grower. 13:30-32.
Davenport
T.L., 1986. Avocado flowering. Hort. Rev. 8:257-289.
Eisikowitch D., and Melamud.
H., 1982, Preliminary experiments for increasing avocado pollination rates (In Hebrew).
Alon Hanotea. 37:19-29.
Ish-Am G., and Eisikowitch
D., 1991. Possible routes of avocado tree pollination by honeybees. Acta Hort.
288:225-233.
Ish-Am G., and Eisikowitch
D., Avocado fruit set is limited by low attractiveness of its flowers to honey
bees. J. Amer. Soc. Hort. Sci. (Submitted for Publication).
Robinson W.S., Nowogrodzki
R., Morse R.A., 1989. The value of honey bees as pollinators of U.S. crops.
Amer. Bee J. 129:411-423, 477-487.
Sokal R.R., and Rohlf F.J.,
1981. Biometry. The Principles and Practices of Statistics in Biological
Research. State Univ. of New York, Stony Brook N.Y. pp. 859.
Vithanage H.I.N.V., 1986.
The role of insects in avocado pollination. In: Williams E.G., Knox R.B. and
Irvine D. (eds.): Pollination'86, Proceedings of a Symposium. p:42-47.
Vithanage H.I.N.V., 1990.
The role of the European honeybee (Apis mellifera L.) in avocado
pollination. J. Hort. Sci. 65:81-86.