Proceedings
of The World Avocado Congress III, 1995 52-56
A
STUDY OF SELF-POLLINATION AND CROSS-POLLINATION IN AVOCADO (PERSEA AMERICANA
MILL.) CV. HASS OF DIFFERENT VARIETIES.
F.J. Gardiazabal and S.
Gandolfo
Facultad de Agronomía. Universidad Católica de Valparaiso Quillota, Chile.
Abstract
The following study was
carried out al "La Palma!' Experimental Center of de Faculty of Agronomy
of de Catholic University of Valparaiso, for the evaluation of the influence of
different avocado varieties as pollinators of Hass. A Hass test field (planted
l0xl0m.) was used, containing sections with different pollinators planted in
1975. Pollinators are distributed in quincunce in the Hass planting, belonging to
the Bacon, Edranol, Hass, Rincon and Zutano varieties. The control was a
section with only Hass trees planted without pollinators. The aim was to
determine for each combination and the control the percentages of
self-pollination and cross-pollination in the descendants of the Hass fruits by
analyzing the origin of parental pollen in fruit embryos using isoenzymes as
genetic markers; thereby enabling a determination of which varieties behave
better as pollinators of Hass under local conditions in the Quillota area.
Electrophoresis test in starch gels at 11% concentrations were carried out
using 14 isoenzymatic systems of GOT-1, GOT-2, PGI-2, PGM-1, PGM-2, LAP-1,
LAP-2, EST-FL, MDIL TPI-2, SKDH-1, SKDH-2, 6-PGD-2, 6-PGD-3 and 6-PGD-4. The
analyzed fruit was collected from October 1994 to January 1995. The results
showed Hass to be weak self- pollinator, being easily displaced by pollen from
other parental plant when found associated to other varieties. The pollinators
which provided the highest cross-pollination results in their combinations were
Zutano, Edranol and Bacon cultivars, presenting no significant differences
between them. Rincon variety proved a weak pollinator for Hass.
1.- Introduction
Avocados have many production
problems, generally related to pollination, biannual production and fruit set
problems, for that reason alternatives need to be found which would allow to
improve or alleviate these issues, thereby attaining larger quantities of
good-quality fruit at harvest.
Pollination problems are due
to the avocado's particular flowering pattern, and despite complete flowers
being present the phenomenon of dicogamia protoginea occurs where the sex
verticils ripen out of time, thereby lessening the chances for self-pollination.
Added to this is the fact of having two flowering patterns, A and B, which open
at different times, and which possibly need to be complementary for adequate
pollination.
For this reason, for a long
time the need to in between trees which are complementary in their flower-type
has been proposed, thereby achieving the maximum pollination and resulting a
high level fruit production. However, it such behavior is not always observed
because is strongly influenced by climatic conditions. It is known that low
temperatures cause delay in blossoming (Sedgley and Grant, 1983), or even the A
and B patterns are not followed (Calvert, 1993).
It is necessary
to evaluate what influence different pollinator trees have on the Hass tree and
analyze whether there is a better pollinating variety for the cv. Hass, than
the Hass itself. For this purpose use was made of one of the orchards belonging
to the Faculty of Agronomy
of the Catholic University of Valparaiso, where there are different fields of
Hass in combination with other varieties.
The results of fruit
production by Hass tress in these different combinations, in studies carried
out several seasons ago, did not show great differences between them. This
indicates that either there is no reason to use pollinators in the case of
Hass, thereby allowing to plant dense orchards of this variety alone,
and producing significant crops, as affirmed by Gazit (1977) and Hodgson
(1947), or, that due to the proximity of other fields, there was probably
significant cross-pollination.
The aim of this study was to
determine the origin of the parental pollen fertilizing the Hass flowers, in
each of the different combinations, through characterization of the
isoenzymes present in the fruit embryo.
2. Material and methods
The study was carried out in
one of the orchards at the "La Palma" Experimental Station at the
Catholic University of Valparaiso, in Quillota Province, Chile (32º 50’
latitude south and 71º 13' longitude west).
This orchard was planted in
1975, with a separation of 5x5 m except for the control Hass which was left
finally at a separation of 10x10m. Later the diagonals were thinned, so at
present the Hass trees are at a l0x10 m separation, with the pollinator at the
center (quincuncial). The fields, planted at random with three repetitions,
have the following combinations:

Each field at
present has 32 trees (except the control field which has only 20), of which 20
correspond to Hass and 12 to the pollinating tree. The sampling material was obtained
by selecting three of the six central Hass trees in each field to avoid, as far
as possible, the influence of the neighboring fields.
Electrophoresis in starch gel
was carried out between the months of October and November 1994, taking fruit
of approximately 12 to 13 months of age. 15 fruits were collected per tree,
choosing 5 large fruits (between 205 and 245g.), 5 medium fruits (173 to 204g.)
and 5 small fruits (150 to 172g.): 45 fruits per field, 13 5 per treatment and
8 10 in total.
Nine enzymes with 15
isoenzymatic systems were analyzed in the samples, in order to be able to
differentiate fruit-parentage as completely as possible. The enzymes which were
analyzed were: GOT-1, GOT-2; PGI-2; PGM-I, PGM-2; LAP-1, LAP-2; EST-FL; MDH;
TPI-2; SKDH-1, SKDH-2; 6-PGO-2, 6-PGO-3 and 6-PGD-4.
3. Results
To determine the parent
supplying the pollen of the progeny, it is the best to use isoenzymes whose
phenotype is homozigote and different between parents, due to the fact that in
this case all progeny can be differentiated. However this situation did not
occur and isoenzymes had to be chosen which had heterozigote phenotypes, with
which only one quarter or half of the descendants could be differentiated, but
nevertheless it was possible to mark out possible parents in certain particular
cases.
A small number of fruits
could not be analyzed, some of large sizes, but mainly small ones whose
embryos, without showing apparent symptoms of being dead or of being altered in
any way, displayed a minimum of intensity in the electrophoresis dyes, or a
total absence of enzymatic activity.
The results, showed a
relatively high number (38%) of cases where it was not possible to identify a
single pollen donor, due to the fact that its gene types did not provide enough
information to permit discrimination from the other possible parents. Ruling
out these doubtful cases could lead to a distortion of the real values of
cross-pollination although in the full study each of the situations was
analyzed. On proving that there are no substantial changes between the two
analyses, we have preferred to give the results considering that when there are
two possible parents the probability of occurrence was distributed equally
between them, 50-50, and in the case of three possible parents the probability
was distributed 1/3 to each one.
Analyzing the percentages of
cross-pollination between treatments, it can be seen that the percentage of
cross-pollination is extremely high in the different fields, with significant
differences between them (Figure 1). These results show that in the fields
there was at least 64.1% cross-pollination, corresponding to the treatment
Hass/Hass, i.e., only 35.9% of the fruit produced by Hass trees were pollinated
with pollen from this same variety.
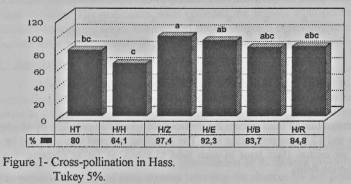
Fields with
pollinators in the quicuncial, such as Rincon, Bacon, Edranol an Zutano,
display significant differences from the Hass/Hass treatment, and indicate
between 84.5% and 97.4% cross-pollination. The control Hass is at an
intermediate point, displaying significant differences only from the
Hass/Zutano treatment.
Avocado orchards in Chile,
including the orchard under study, are managed with bees, with between 3 and 6
hives placed in each hectare. It is, therefore, not surprising to find these results,
bearing in mind the large radius of action these pollinating insects exert in
free-pollination orchards, and taking into account the scant separation between
the different fields.
Analysis
of cross-pollination between each treatment (Table 1), shows that there is an
interaction between different pollen-donating parents. In the treatments
Hass/Control, Hass/Hass, and Hass/Rincon, the Zutano, Edranol and Bacon parent-
trees participated strongly. On the other hand when these parent trees are
present in the fields they themselves dominate strongly.
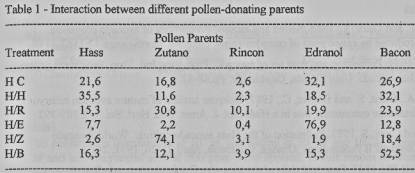
The weakness of
Hass as a self-pollinator has already been reported by Guil and Gazit
(1992).
There may be several explanations: first, Hass may have some problems with its
pollen which makes self-pollination difficult; second, climatic conditions may
not favor a good overlap of blossom states, thereby favoring cross-pollination;
third, the genetic material which the Hass plant passes on to its descendants
is in a disadvantage compared with other pollen donors, as far as it allows a
high survival rate of small fruits.
The percentage of
cross-pollination for each treatment, and by sizes, shows that in the total
number of fruits, the Zutano, Edranol and Bacon varieties are statistically the
same, but they differ from Hass, which in its turn differs from Rincon (Table
2).
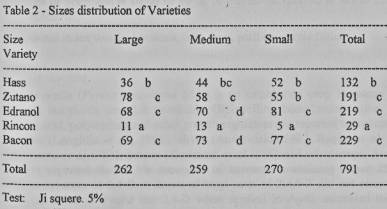
It can be seen that Rincon
has a small share as much in the different treatments as in sizes. It should
not be forgotten that this is the only cultivar which has a type-A flower, like
the Hass variety which is the one to be pollinated. However, if Hass did
self-pollinate perhaps this variety's problem lies in the quality of pollen, as
suggested by Degani and Gazit (1984).
All
the other varieties have type-B flowers, which means that their blossom states
are complementary with Hass, and perhaps with better pollen
"quality", possibly implying a greater fruit set and a higher
survival rate of fruits up to maturity. For that reason it can be deduced that
the genotype of the embryo plays a very important role in
differential-selection for fruit survival.
References
Degani, C. and Gazit, S., 1984. Selfed and crossed
proportions of avocado progenies
produced by caged pairs of complementary cultivars.
HortScience 12:1187-1188.
Gazit, S. 1977. Pollination
and fruit set of avocado. Proc. First Intl. Trop. Fruit short course: The
avocado. Univ Florida, Gainesville. Pp. 88-92.
Goldring, A., Gazit, S. and
Degani, C., 1987. Isozyme analysis of mature avocado embryos to determine outcrossing
rate in a Hass plot. J. Amer. Soc. Hort. Sci. 112:3 89-3 92.
Guil, I. and
Gazit, S. 1992. Pollination of the Hass
avocado cultivar. World Avocado
Congress
II Proceedings. Orange, California, April
21-26, 1991. 241 p.
Hodgson, R.W., 1947. Bearing habits of the avocado.
California Avocado Society
Yearbook p:35-39.
Sedgley, M. and Grant, W.J., 1983. Effect of low
temperatures during flowering on floral
cycle and pollen tube growth in nine avocado
cultivars. Scientia. Horticulturae 18:207-213.
Torres, A.M. and Bergh, B.O., 1980. Fruit and leaf
isozymes as genetic markers in
avocado. J. Amer. Soc. Hort.
Sci. 105:614-619.
Vrecenar-Gadus, M. and Ellstrand, N.C., 1984.
Independent assortment of four isozyme loci in the Bacon avocado (Persea
americana Mill.). Cal Avoc. Soc.Yearbook 68:173-177.