Proceedings
of The World Avocado Congress III, 1995 172 - 180
INFLUENCE OF AUTUMN FERTELIZATION WITH NITROGEN AND PHOSPHORUS ON ROOT ACTIVITY AND DEVELOPMENT IN AVOCADO
Leo Winer
Baruch
Reuveni Yoel Bar
Ministry
of Agriculture Kibbutz Nir-Eliau 45845 The
Hebrew University of Jerusalem
Ranaana 43208
Israel Rehovot 76100
Israel Israel
Jerry
H. Haas and Aida Zveibil
Dept. Plant Pathology
Agricultural Research Organization
Bet-Dagan 50250
Israel
Abstract
Autumn fertilization with nitrogen
(N) and phosphorus (P) for eight years increased yield by 50% in avocado (Persea
americana). The supplementary NP-treatment was compared with normal spring
and summer fertilization alone. The yield increase was accompanied by increased
root mass and activity in the spring, as monitored by increased white root tip
density. The roots of NP-treated trees had higher levels of calcium (Ca) and
endomycorrhiza.
The concentration of P in
roots was higher in NP-treated trees, while no differences were found in the
concentration in leaves. The mineral composition was different on roots of
different diameters. In thin roots (<1-mm-wide) of NP-treated trees, the P
concentration was twice as high and there was a change in the quantitative
relationship between the cations. NP treatment decreased the concentration of
potassium and magnesium and increased the concentration of Ca. Also, in wider
roots, there was a change in element concentration with trends similar to those
in thin roots although not always as great as in thin roots. The concentration
of microelements in roots was not affected by NP autumn fertilization.
Further work must be done to
determine the element concentration in roots during the growth season to verify
the possibility of using these data as a measure of the nutrient status of the
tree.
Additional index words: yield, mycorrhiza, Persea
americana, nutrition, root morphology
Introduction
Root morphology and
root-shoot relationships are affected by mineral nutrition (Aung, 1974;
Hackett, 1968). In general, nitrogen (N) causes an increase in root mass and
number of root hairs, phosphorus (P) and potassium (K) cause and increase in
ramification of the roots, and calcium (Ca) improves root hardiness (Vogt et
al., 1991).
Sub-optimal
mineral nutrition, especially P and N, often results in an increase in
root/shoot ratio (Alberda, 1965; Barley, 1970; Bourna, 1967; Taylor et al.,
1976). Sometimes elongation of the main root occurs at the expense of the
lateral roots at sub- optimal K (Jensen, 1982) and N (Clement et al., 1979)
levels.
A different kind of root
morphology modification can be found when only a limited volume of roots of
plants with mineral deficiencies are fertilized (Russel, 1977). In this case
there is an increase in number of lateral roots, especially with N and P
fertilization (Drew et al., 1973; Drew et al., 1975). With K there is also a
general increase in the number of lateral roots, and not only in limited
fertilized volume (Drew et al., 1975). The mineral nutrition, especially of N, and
P, can also increase the length and density of the thin roots.
The form of N supplied to the
root can influence the growth and viability of the root. Ammonium leads to a
lower root turnover and biomass, and nitrate leads to a higher root turnover
and biomass (Aber et al., 1985; Smucker, 1984).
Root mass is in
part controlled by the photosynthate source and competition between sinks that
determine the carbohydrate distribution. There is a slow turnover of the thick
roots (Vogt et al., 1991) and a fast turnover of the thin roots (Cox et al.,
1978; Reynolds, 1970; Vogt et al., 1991).
Thin roots are generally
associated with mycorrhizal fungi (Vogt et al., 1981) that can influence their
longevity. In forest trees the lack of mycorrhiza can shorten root longevity to
a few weeks; with mycorrhiza the roots can remain active for some years (Vogt
et al., 1991).
In a previous
work we reported an highly significant yield increase in avocado by autumn
fertilization with N and P (Reuveni et al., 1989). There was an increase in
yield despite the similar concentrations of N and P as determined by standard
leaf samples of NP-treated and control trees. The highest yield in NP-treated
trees was accompanied by greater leaf number during the spring, and reduced
leaf abscission during fruiting season (Reuveni et al., 1989). An occurrence of
extremely hot winds in May 1988, which caused a large decrease in Israel
avocado yield, lead to a more rapid old-leaf drop in control trees compared
with NP-treated trees (Reuveni et al., 1989).
Preliminary observations on
the rapidity of root renewal in the spring by NP-treated trees in the present
work, lead us to hypothesize that the increased yield is a result of the
nutrient treatment effect on root growth and morphology. In the present work we
determined the effect of autumn fertilization with N and P on the element
composition of roots and the rapidity of root renewal in the spring.
Materials
and methods
An avocado (Persea
americana Mill.) plantation growing on sandy-loam soil, pH 7, in Kibbutz
Nir-Eliahu in the Coastal Plain of Israel was selected. The trial was initiated
in November 1985 when the plantation was 7-years-old. The trees were cv. 'Hass'
on Nabal stock. A group of thirty trees was selected for the trial. Ten
completely randomized blocks of 3 consecutive trees in a row were established.
Five blocks were subjected to an NP-fertilizer treatment during November of
each year. Five blocks served as non-treated controls.
Standard horticultural
treatments. The
irrigation was effected with mini-jets twice a week. During spring and summer
the plantation fertilizer was distributed through the irrigation system. A
total of 25 kg N/ 1000 m2 was applied each season. Potassium
was applied only during 1988 and 1989 at a rate of 25 kg K/ 1000 M2 per season.
Autumn fertilization
treatment The
NP-fertilizer treatments were applied through the irrigation system twice each
year during two successive irrigations of 6 mm. An "8-26-0"
fertilizer was prepared with ammonium nitrate and phosphoric acid and injected
into the irrigation water; the mean amount applied each autumn during the eight
years of experiment was 2 kg N and 6.5 kg P2O5 / 1000 M2 per season.
Root and soil sample. Soil samples were taken in
March each year with a 0.5-liter- volume metal tube (8-cm-diarneter X
10-cm-length). The samples came from the irrigated region under the tree canopy
at a distance of 40 to 50 cm from a mini-jet. Leaf debris was cleared away prior
to taking the core which contained soil and roots. Six to 8 cores were sampled
from the central tree of each replicate. The soil was separated from the roots
by sieving through a 2 mm screen taking care to prevent loss of small roots.
The screened soil was taken for chemical analysis. Roots were used for
determination of root number, mass and mineral composition.
Determination of root dry
weight and mineral composition. The total number of white-colored root tips visible
during the spring flush served as the measure of root activity. Because so many
roots tips were counted at the first counting in May 1991, we decided to
determine root activity earlier (March) in the following years. After counting
the root tips, all the roots were classified and separated according to their
diameter and color: white roots (a) 0 to 0.9 mm, (b) 1 to 1.9 mm, (c) 2 to 3
mm, and dark roots.
After washing
the roots with distilled water they were prepared for mineral determination.
Roots were placed in sulfuric acid 10% (v/v) for a few seconds, rinsed with
distilled water and dried at 60 ºC for a few days (until there was no change in
sample weight). The light colored roots were ground in a Wiley mill and mineral
composition was determined in the Plant Nutrition Laboratory of the Faculty of
Agriculture, Rehovot.
Association
of thin roots with mycorrhizal fungus. Root samples for evaluating the percent of root
colonized by mycorrhizal fungi were collected on 30 July 1992, in a similar way
to that of the spring root sampling. A composite sample from the several soil
cores around each central-tree was screened, washed under running water, and
the remaining light-colored roots were randomly separated into 10 g aliquots.
The preparation of the roots
for mycorrhizal determination was done as described by Haas et al. (1986). The
roots were cleared in 2% KOH at 95 ºC rinsed in 1% HCl and stained with
Trypan-Blue.
The percent of roots
colonized with endomycorrhizal fungus was estimated by the visual evaluation
method as described by Giovannetti et al. (1980).
Leaf analysis. In October 1993, 15 mature
leaves were sampled from the central tree of each block and nutrient analysis
was performed in the Plant Nutrition Laboratory of the Faculty of Agriculture,
Rehovot.
Yield Avocado fruits from the three
trees of each block were collected separately and weighed during each year of
the trial.
Statistical analysis. Analysis of variance was
performed separately on the data from each sampling. Means separation was by
F-test or Duncan's Multiple Range Test, as appropriate.
Results
Soil and leaf analysis. In the spring of 1992, seven
years after the start of autumn NP- treatments, there was no significant
fertilizer effect on the N or K concentrations or electrical conductivity in
the soil. There was 3.0 mg NI kg soil, 4.3-4.6 mg K/ kg and conductivity was
0.35-0.36 dS/ in. The available P under NP-treated trees was 27 mg/ kg and that
under control trees was 13 mg/ kg.
Concentrations of N, P, K and
Ca were similar in leaves of NP-treated and control trees sampled in October
1993, the last year of the trial. On a dry weight basis the mean of NP-treated
and control roots was 1.82 and 1.88 % N, 0.196 and 0.188 % P, 1.01 and 1.06 %
K, and 2. 10 and 2.17 % Ca, respectively.
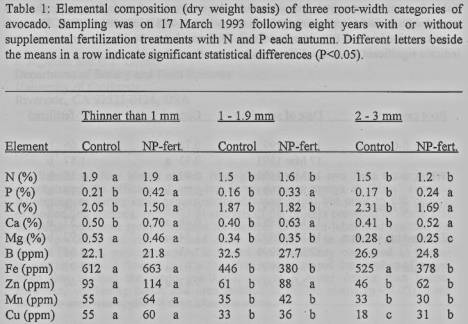
Mineral composition of the
roots. The
mineral composition of the thin (up to 3-mm- wide), light-colored roots sampled
in March 1992 and 1993 was similar in both years and only the 1993 results are
presented (table 1). The concentration of some elements differed in roots of
different width. The N, Mg, Zn, Mn and Cu concentrations were higher in the
thinnest roots (<I-mm width) and decreased with the increase in root width.
The autumn NP-fertilization greatly increased the concentration of P and Ca in
the roots (P<0.01), especially those up to 2-mm diameter. The autumn
NP-fertilization caused a decrease in K concentration of the thinnest roots
(<1-mm width) and of the widest roots (2 to 3-mm width). There were no
significant differences in the concentration of the other elements measured.
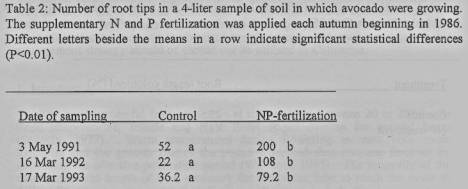
Root mass and activity. The number of root tips in
the 4-liter soil collected served as a measure of root activity in spring. The
autumn NP-fertilization stimulated earlier and increased root activity. In the
mid-March and early May there were two to five times more root tips in
NP-treated trees than in control trees (table 2).
The dry weight
of roots in the sample was considered a measure of the total tree root mass.
The autumn NP-fertilization resulted in a doubling or tripling in the small
root mass (table 3). The weight of larger, dark roots was increased by 50% by
the fertilization treatment (table 3).
Yield The average yield of the
8-harvests from 1986 to 1993 was significantly higher in NP-treated trees.
There were 29.5 kg fruit/ tree/ year in control trees and 44.2 kg in treated
plants.
Mycorrhizal symbiosis. Autumn NP-fertilization
caused an increase in the association of thin roots with mycorrhizal fungi .
The mycorrhizal fungus colonization was doubled by the NP-treatment (table 4).
Discussion
Supplementary fertilization
with N and P for an eight-year-period from 1986-1993 caused a 50% increase in
mean yield of avocado. This increase was recorded despite the lack of evidence
of N or P deficiency in leaves, based on the levels generally accepted in
Israel (2% N and 0. 12% P, dry weight basis).
The K concentration was higher in leaves of control
trees during the first years of the trial (1986 to 1989) (Reuveni et al.,
1989). This difference in K concentration was annulled by fertilizing both
treatments with K in 1988 and 1989.
The increased yield following autumn NP-fertilization
challenges the validity of the standard leaf analysis recommendations for
sufficiency of these two elements. In a previous report (Reuveni et al., 1989)
we suggested the need for selecting a different plant tissue for nutrient
status analysis of avocado. This is one consideration that lead us to examine
the mineral composition of roots and to determine the kinetics of the changes
in mineral composition of leaves and flowers during spring (Yoel Bar and Leo
Winer, paper in preparation).
In the present work we found differences in nutrient
concentration of roots depending upon the root width. The most important
differences in the nutrient concentration in roots following autumn NP-fertilization
were found in the thin roots (<1-mm-width). The P concentration in roots was
increased and K concentration decreased by autumn NP fertilization.
More work must be done to determine the time-course of changes in element concentrations in roots during the season, before these data can be employed as additional parameters for determining the mineral nutrition status of avocado.
One of the important findings of the present work is the effect of autumn NP- fertilization on the greater intensity of root activity in the spring and, after eight years of the trial, a very significant increase in total root mass. The higher total root mass may be the result of the prolonged root longevity. Many factors are known to influence root development and longevity. Brown et al. (1984) thought that soil parameters are paramount to genetic parameters in determining the rate of root turnover. This would lead to more intensive root development in the parts of the soil mass with less water or nutrient deficiency and to greater nutrient uptake by the roots (Eissenstat et al., 1988; St. John et al., 1983). This, in contrast with more rapid aging and root death in the adequately watered and fertilized parts of the soil.
The autumn NP-fertilization
increased the symbiosis of thin roots and mycorrhizal fungi. The mycorrhizal
symbiosis is known to positively affect the longevity of roots (Vogt et al.,
1991), and cause increased concentrations of Ca, an element known to strengthen
roots (Vogt et al., 1991). An important effect of mycorrhiza is to improve the
transfer of available-P to roots and this was evident in our results. High
levels of P in roots can inhibit mycorrhizal development but not at the levels
found in the treated-tree roots.
We postulate that the
increased root growth and avocado yield was largely due to the greater
absorption of P in treated trees and that the mycorrhizal fungi contributed
significantly to this absorption.
References
Aber, J.D., Melillo, J.M., Nadelhoffer,
K.L., McClaugherty, C.A,. and Pastor, J., 1985. Fine root turnover in forest
ecosystems in relation to quantity and form of nitrogen availability: a
comparison of two methods. Oecologia 66:317-321.
Alberda, Th., 1965. Influence
of temperature, light intensity, nitrate concentration on dry matter production
and chemical composition of Lolium perenne L., Neth. J. Agric. Sci.
13:335-360.
Aung, H.L., 1974. Root-shoot
relationships. In: The Plant Root and its Environment. E.W. Carson, Ed.,
University Press of Virginia, Charlottesville, pp. 29-61.
Barley, K.P., 1970. The
configuration of the root system in relation to nutrient uptake. Adv. Agronomy
22:159-201.
Bouma, D., 1967. Nutrient
uptake and distribution in subterranean clover during recovery from nutritional
stress. 1. Experiments with P. Aust. J. Biol. Sci. 20:601-612.
Brown, D.A. and Scott, H.D.,
1984. Dependence of crop growth and yield on root development and activity. Ln:
Roots, Nutrient and Water Influx, and Plant Growth, Publication No. 49 *
American Society of Agronomy, Madison, Wis., pp. 10 1 - 13 6.
Cox, T.L., Harris, W.P.,
Ausmus, B.S. and Edwards, HT., 1978. The role of roots in biogeochemical cycles
in an eastern deciduous forest. Pedologia 18:264-271.
Clement, C.R., Jones, L.H.P.
and Hopper, M.J., 1979. Uptake of nitrogen from flowing nutrient solution:
Effect of terminated and intermittent nitrate supplies. In: Nitrogen
Assimilation of Plants. (E.J. Hewitt, C.V. Cuttings, Eds.). Academic Press,
London. pp. 123-133.
Drew, M.C., and Saker, L.R.,
1975. Nutrient supply and the growth of the seminal root system in barley. 11.
Localized, compensatory increases in lateral root growth and rates of nitrate
uptake when nitrate supply is restricted to only part of the root system. J.
Exp. Bot. 26:79-90.
Drew, M.C., Saker, L.R. and
Ashley, T.W., 1973. Nutrient supply and the growth of the seminal root system
in barley. I. The effect of nitrate concentration on growth axes and laterals.
J. Exp. Bot. 24:1189-1202.
Eissenstat, D.M., and Caldwell,
M.M., 1988. Seasonal timing of root growth in favorable microsites. Ecology
69:870-873.
Giovannetti, M. and Mosse,
B., 1980. An evaluation of techniques for measuring vesicular arbuscular
mycorrhizal infection in roots. New Phytol. 84:489-500.
Haas, J.H., Bar-Tal, A.,
Bar-Yosef, B., and Krikun, J., 1986. Nutrient availability effects on vesicular
arbuscular mycorrhizal bell pepper (Capsicum annuum L.) seedlings and
transplants. Ann. App. Biol. 108:171-179.
Hackett, C., 1968. A study of
the root system of barley. 1. Effects of nutrition on two varieties. New
Phytol. 67:287-299.
Jensen, P., 1982. Effects of
interrupted K supply on growth and uptake of K, Ca, Mg, and Na in spring wheat.
Physiol. Plant. 56:259-265.
Orlov, A.Y., 1957.
Observations on absorbing roots of spruce (Picea excelsa Link) in
natural conditions. Bot. Zh. SSSR 42:1172-118 1.
Reuveni, B., Winer, L., and
Shnior, 1., 1989. Autumn fertilization for yield increase in avocado. Alon
Hanotea 3:191-198 (in Hebrew).
Reynolds, E.R.C., 1970. Root
distribution and the cause of its spatial variability in Pseudotsuga
tasifiblia (Poir.] Brit. Plant Soil 32:501-517.
Reynolds, E.R.C., 1975. Tree
rootlets and their distribution. In: The Development and Function of
Roots (J.G. Torrey, D.T. Clarkson, Eds.). Academic Press, London, pp. 163-177.
Russel, R.S., 1977. Plant
Root Systems. McGraw-Hill, New York. pp. 53-61, 82-89. Smucker, A.J.M., 1984.
Carbon utilization and losses by plant root systems. In: Roots, Nutrient
and Water Influx, and Plant Growth, Special Publication No. 49. American
Society of Agronomy, Madison, Wis., pp. 27-46.
St. John, T.V., Coleman,
D.C., and Reid, C.P.P., 1983. Growth and spatial distribution of
nutrient-absorbing organs: selective exploitation of soil heterogeneity. Plant
Soil 71:487-493.
Taylor, B.K., and Goubran,
F.H., 1976. Effects of phosphate and pH stress on the growth and function of
apple roots. Plant Soil 44:49-162.
Vogt, K.A., Bloomfield, J.,
1991. The root turnover and senescence. In: Plant Roots, the Hidden
Half. (Y. Waisel, A. Eshel, U. Kafkafi, Eds.). Marcel Dekker, New York. pp.
287-306.
Vogt, K.A., Edmonds, R. L.,
and Grier, C.C., 1981. Seasonal changes in biomass and vertical distribution of
mycorrhizal and fibrous-textured conifer fine roots in 23 and 180-year-old subalpineAbiesamabilis
stand. Can. J. For. Res. 11:223-229.